


Information
Author: Jenny Volstorf
Version: B | 1.1Published: 2022-01-22
Please note: This part of the profile is currently being revised.
1 Remarks
1.1 General remarks
Escapees and consequences: negative or at most unpredictable for the local ecosystemPreferences: opportunistic
1.2 Other remarks
No data found yet.2 Ethograms
In the farm or lab: on daily rhythm, communication, social behaviour, coping styles
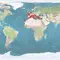
3 Distribution
Natural distribution: eastern Atlantic, Mediterranean
Introduced: eastern Atlantic, Pacific ocean
4 Natural co-existence
5 Substrate and/or shelter
5.1 Substrate
Substrate range, substrate preference: opportunistic – reported from seagrass beds, salt marsh creeks, muddy and rocky bottoms5.2 Shelter or cover
Shelter or cover preference: sand for cover during night (further research needed)6 Food, foraging, hunting, feeding
6.1 Trophic level and general considerations on food needs
Trophic level: 3.7Impacts of feed fishery: contributes to overfishing, challenges animal welfare
6.2 Food items
Food items, food preference: mainly carnivorous, increasing prey size with increasing ageFeed enrichment and stress tolerance: direct effect of arachidonic acid, probiotic Pdp11, and vitamin E
6.3 Feeding behaviour
Feeding style, foraging mode: benthicFeed delivery and stress: unpredicted schedule increases stress and swimming activity
Feed delivery and growth: inverted u-shaped relation (further research needed)
Food competition and stress: direct effect (further research needed)
Food competition and growth: inverse effect for subordinate individuals
Effects on feeding: direct relation with temperature until 30 °C
For feeding and...
...vision → D2,
...olfaction → D3,
...establishing hierarchy → D4,
...dominance → D5.
7 Photoperiod
7.1 Daily rhythm
Daily rhythm: diurnal, nocturnal when solitary or in small groupPhotoperiod and growth: direct relation (further research needed)
7.2 Light intensity
No data found yet.7.3 Light colour
Light colour and stress: red light is disadvantageous (further research needed)8 Water parameters
8.1 Water temperature
Standard temperature range, temperature preference: 11-30 °CTemperature and stress: abnormalities and decreasing survival <16 °C and >22 °C (further research needed)
Temperature and growth: optimally 25 °C, the less fluctuations the better (further research needed)
8.2 Oxygen
No data found yet.8.3 Salinity
Salinity tolerance, standard salinity range: probably euryhaline (further research needed)Salinity and stress: direct relation between salinity tolerance and age (further research needed)
Salinity and growth: salinity of 18-28‰ benefits growth in fry
8.4 pH
No data found yet.8.5 Turbidity
No data found yet.8.6 Water hardness
No data found yet.8.7 NO4
No data found yet.8.8 Other
No data found yet.9 Swimming
9.1 Swimming type, swimming mode
Swimming type, swimming mode: carangiform9.2 Swimming speed
Swimming speed: 0.5-3.5 body lengths/s (adults), relatively decreasing with body length, decreasing above and below 25 °C (further research needed)9.3 Home range
Home range: up to 4 km (further research needed), site fidelity9.4 Depth
Depth range, depth preference: 0-30 m, seldomly until 150 m9.5 Migration
Migration type: amphidromous10 Growth
10.1 Ontogenetic development
Mature egg: 0.8-1.0 mmLarvae: hatching to 2-5 days, 2.5+ mm
Fry: beginning of exogenous feeding, 4-160 days, 5.5-71.5 mm
Juveniles, sexual maturity: fully developed (160 days) to beginning of maturity (first or second year), 2-32.6 cm, 0.02-0.3 kg
Maturation and manipulation: advanced photoperiod delays maturation (further research needed)
Adults: 1-12 years, 20.5-70 cm, 1.5-3.4 kg
10.2 Sexual conversion
Sexual conversion: juveniles develop first into males at 2-3 years, 10-30 cm, then turn into females at 2-3 years, 15-45 cmSex and manipulation: adding younger individuals or females influences male:female ratio of the broodstock (further research needed)
10.3 Sex ratio
No data found yet.10.4 Effects on growth
Growth rate: 100-200 g/year during April-OctoberGrowth and other factors: offshore cages and music benefit growth (further research needed)
For growth and...
...feed delivery → D18,
...food competition → D19,
...PHOTOPERIOD → D20,
...water temperature → D21,
...salinity → D22,
...stocking density → D23.
10.5 Deformities and malformations
Deformities and malformations: skeletal deformities and morphological malformations in 7.8-100% of individualsHypotheses regarding causes for deformities: bacteria, uninflated swim bladder, missing natural selection, physical stress, inheritance
11 Reproduction
11.1 Nest building
Nest building: none11.2 Attraction, courtship, mating
Attraction: increasing pigmentation in both sexes July-October (further research needed)11.3 Spawning
Spawning conditions: usually October-MarchMale:female ratio resulting in spawning, composition of the broodstock: small groups of mixed sex (further research needed)
Spawning sequence: 10-30 min (further research needed)
11.4 Fecundity
Female fecundity: 4,100-80,000 eggs daily for 4-100 days (further research needed)Effects on fecundity: direct effect of full moon
Fecundity and manipulation: increasing temperature and implanting GnRHa delivery systems increases fecundity
11.5 Brood care, breeding
Breeding type: sea spawner, larvae migrate to nursery grounds (lagoons, estuaries)12 Senses
12.1 Vision
Importance of vision: foraging (further research needed)Substrate colour preference: blue and red-brown (further research needed)
12.2 Olfaction (and taste, if present)
Olfactory spectrum (and gustatory, if present): intestinal, seminal, and egg fluid, urine, amino acids (further research needed)Importance of olfaction: foraging (further research needed)
12.3 Hearing
Hearing type, hearing spectrum: hearing generalistImportance of hearing: orientation (futher research needed)
Noise and stress: mixed effects (further research needed)
For music and growth → D27.
12.4 Touch, mechanical sensing
No data found yet.12.5 Lateral line
Importance of lateral line: detecting local water movements and low frequency waves (further research needed)12.6 Electrical sensing
No data found yet.12.7 Nociception, pain sensing
No data found yet.12.8 Other
No data found yet.13 Communication
13.1 Visual
Signalling aggression: darker body colouration, erect dorsal fin (further research needed)13.2 Chemical
No data found yet.13.3 Acoustic
Sound production: via contracting sonic muscles around the swim bladder13.4 Mechanical
No data found yet.13.5 Electrical
No data found yet.13.6 Other
No data found yet.14 Social behaviour
14.1 Spatial organisation
Aggregation type: solitary or in small to large aggregationsStocking density and stress: direct relation from ca 22 kg/m3 on (further research needed)
Stocking density and growth: inverse relation (further research needed)
14.2 Social organisation
Social organisation type: linear hierarchy (when in small groups)Features of dominance: occupy and patrol best feeding sites, more aggressive than subordinates
Features of subordination: hardly move, stay away from food
14.3 Exploitation
No data found yet.14.4 Facilitation
No data found yet.14.5 Aggression
Aggression and size-grading: chasing, nipping regardless of size-grading (further research needed)Effects on aggression: no sex bias (further research needed)
For aggression and...
...substrate colour → D29,
...dominance → D5,
...learning → D30,
...coping styles → D31.
14.6 Territoriality
No data found yet.15 Cognitive abilities
15.1 Learning
Learning and aggression: handling experience might increase aggression (further research needed)15.2 Memory
No data found yet.15.3 Problem solving, creativity, planning, intelligence
No data found yet.15.4 Other
No data found yet.16 Personality, coping styles
17 Emotion-like states
17.1 Joy
No data found yet.17.2 Relaxation
No data found yet.17.3 Sadness
No data found yet.17.4 Fear
No data found yet.18 Self-concept, self-recognition
19 Reactions to husbandry
19.1 Stereotypical and vacuum activities
No data found yet.19.2 Acute stress
Handling: stressfulFor acute stress and...
...food competition → D34,
...light colour → D35,
...salinity → D26,
...noise → D36.
19.3 Chronic stress
For chronic stress and......feed enrichment → D37,
...feed delivery → D38,
...water temperature → D24,
...noise → D36
...stocking density → D39,
...dominance → D5,
...subordination → D32.
19.4 Stunning reactions
Stunning rules: fast, effective, safeStunning methods: percussive and electrical stunning most effective
19.4.3 Stunning methods and stress
Glossary
AGGRESSIVENESS = agonistic reactions towards conspecifics. Tests: mirror image, social interaction/diadic encounters 136.
FARM = setting in farming environment or under conditions simulating farming environment in terms of size of facility or number of individuals
FOOD CONVERSION RATIO = (food offered / weight gained)
FRY = larvae from external feeding on
GENERALIST = Generalists detect a narrow bandwidth of sound frequencies (<50-500 Hz, 1,500 Hz max.). High hearing threshold = cannot detect quieter sounds. Typically no swim bladder or no attachment of the swim bladder to the inner ear. Live in loud environments (rivers) 120121.
JUVENILES = fully developed but immature individuals
LAB = setting in laboratory environment
LARVAE = hatching to mouth opening
MILLIARD = 1,000,000,000 46 47
PHOTOPERIOD = duration of daylight
TOTAL LENGTH = from snout to tip of caudal fin as compared to fork length (which measures from snout to fork of caudal fin) or standard length (from head to base of tail fin) or body length (from the base of the eye notch to the posterior end of the telson) 90
WILD = setting in the wild
Bibliography
2 Alves, Filipe M. A., and Catarina M. A. Alves. 2002. Two new records of seabreams (Pisces:Sparidae) from the Madeira Archipelago.
3 Dimitriou, Evagelos, George Katselis, Dimitrios K Moutopoulos, Constantin Akovitiotis, and Constantin Koutsikopoulos. 2007. Possible influence of reared gilthead sea bream (Sparus aurata, L.) on wild stocks in the area of the Messolonghi lagoon (Ionian Sea, Greece). Aquaculture Research 38: 398–408. https://doi.org/10.1111/j.1365-2109.2007.01681.x.
4 Arechavala-Lopez, P., I. Uglem, D. Fernandez-Jover, J. T. Bayle-Sempere, and P. Sanchez-Jerez. 2012. Post-escape dispersion of farmed seabream (Sparus aurata L.) and recaptures by local fisheries in the Western Mediterranean Sea. Fisheries Research 121–122: 126–135. https://doi.org/10.1016/j.fishres.2012.02.003.
5 Palma, J., J.a. Alarcon, C. Alvarez, E. Zouros, A. Magoulas, and J.p. Andrade. 2001. Developmental stability and genetic heterozygosity in wild and cultured stocks of gilthead sea bream (Sparus aurata). Journal of the Marine Biological Association of the United Kingdom 81: 283–288. https://doi.org/10.1017/S0025315401003757.
6 Magoulas, Antonis. 2002. Genetic considerations in the introduction of aquacultured fish to natural ecosystems. In Aquaculture Challenges in Asia After the Bangkok Declaration on Sustainable Aquaculture The next step, 79–85. Beijing, China.
7 Alarcón, J. A., A. Magoulas, T. Georgakopoulos, E. Zouros, and M. C. Alvarez. 2004. Genetic comparison of wild and cultivated European populations of the gilthead sea bream (Sparus aurata). Aquaculture 230: 65–80. https://doi.org/10.1016/S0044-8486(03)00434-4.
8 Šegvić-Bubić, Tanja, Ivana Lepen, Željka Trumbić, Jelena Ljubković, Davorka Sutlović, Sanja Matić-Skoko, Leon Grubišić, Branko Glamuzina, and Ivona Mladineo. 2011. Population genetic structure of reared and wild gilthead sea bream (Sparus aurata) in the Adriatic Sea inferred with microsatellite loci. Aquaculture 318: 309–315. https://doi.org/10.1016/j.aquaculture.2011.06.007.
9 Jobling, Malcolm, and Stefano Peruzzi. 2010. Seabreams and Porgies (Family: Sparidae). In Finfish Aquaculture Diversification, ed. Nathalie R. Le Francois, Malcolm Jobling, Chris Carter, and Pierre Blier, 361–373. CABI.
10 Abecasis, David, and Karim Erzini. 2008. Site fidelity and movements of gilthead sea bream (Sparus aurata) in a coastal lagoon (Ria Formosa, Portugal). Estuarine, Coastal and Shelf Science 79: 758–763. https://doi.org/10.1016/j.ecss.2008.06.019.
11 Bauchot, M.-L., J.-C. Hureau, and J. C. Miguel. 1981. Sparidae. In FAO species identification sheets for fishery purposes. Eastern Central Atlantic., ed. W. Fischer, G. Bianchi, and W. B. Scott. Vol. 4. Rome: FAO.
12 Bégout, Marie-Laure, and Jean-Paul Lagardére. 1995. An acoustic telemetry study of seabream (Sparus aurata L.): first results on activity rhythm, effects of environmental variables and space utilization. Hydrobiologia 300–301: 417–423. https://doi.org/10.1007/BF00024483.
13 Arabaci, Muhammed, Yasin Yilmaz, Saltuk Bugrahan Ceyhun, Orhan Erdogan, Hakan Galip Dorlay, Ibrahim Diler, Suleyman Akhan, et al. 2010. A Review on Population Characteristics of Gilthead Seabream (Sparus aurata). Journal of Animal and Veterinary Advances 9: 976–981. https://doi.org/10.3923/javaa.2010.976.981.
14 Paspatis, M., D. Maragoudaki, and M. Kentouri. 2000. Self-feeding activity patterns in gilthead sea bream (Sparus aurata), red porgy (Pagrus pagrus) and their reciprocal hybrids. Aquaculture 190: 389–401. https://doi.org/10.1016/S0044-8486(00)00409-9.
15 Velázquez, M., S. Zamora, and F. J. Martínez. 2004. Influence of environmental conditions on demand-feeding behaviour of gilthead seabream (Sparus aurata). Journal of Applied Ichthyology 20: 536–541. https://doi.org/10.1111/j.1439-0426.2004.00613.x.
16 Basaran, Fatih, Huseyin Ozbilgin, and Yeliz Doganyilmaz Ozbilgin. 2007. Comparison of the swimming performance of farmed and wild gilthead sea bream, Sparus aurata. Aquaculture Research 38: 452–456. https://doi.org/10.1111/j.1365-2109.2007.01670.x.
17 Koumoundouros, G., C. Ashton, G. Xenikoudakis, I. Giopanou, E. Georgakopoulou, and N. Stickland. 2009. Ontogenetic differentiation of swimming performance in Gilthead seabream (Sparus aurata, Linnaeus 1758) during metamorphosis. Journal of Experimental Marine Biology and Ecology 370: 75–81. https://doi.org/10.1016/j.jembe.2008.12.001.
18 Kouzoupis, S., and P. Papadakis. 2010. Modeling the swimbladder sound producing mechanism of the Gilthead seabream (Sparus aurata). In . Istanbul.
19 Kouzoupis, S., P. Papadakis, V. Boura, I. Xezonakis, and M. Kentouri. 2007. Preliminary investigation on sound production by two fish species: Sparus Aurata and Dicentrarchus Labrax. In Proceedings of the Institute of Acoustics, 29:225–232. Holywell Park, Loughborough University, UK.
20 Karplus, I., D. Popper, and O. Goldan. 2000. The effect of food competition and relative size of group members on growth of juvenile gilthead sea bream, Sparus aurata. Fish Physiology and Biochemistry 22: 119–123. https://doi.org/10.1023/A:1007885525902.
21 Goldan, Oded, Dan Popper, and IIan Karplus. 2003. Food Competition In Small Groups Of Juvenile Gilthead Sea Bream (Sparus Aurata). The Israeli Journal of Aquaculture - Bamidgeh 55: 94–106.
22 Montero, D., G. Lalumera, M. S. Izquierdo, M. J. Caballero, M. Saroglia, and L. Tort. 2009. Establishment of dominance relationships in gilthead sea bream Sparus aurata juveniles during feeding: effects on feeding behaviour, feed utilization and fish health. Journal of Fish Biology 74: 790–805. https://doi.org/10.1111/j.1095-8649.2008.02161.x.
23 Tandler, A., M. Har’el, M. Wilks, A. Levinson, L. Brickell, S. Christie, E. Avital, and Y. Barr. 1989. Effect of environmental temperature on survival, growth and population structure in the mass rearing of the gilthead seabream, Sparus aurata. Aquaculture 78: 277–284. https://doi.org/10.1016/0044-8486(89)90105-1.
24 Castanheira, Maria Filipa, Marcelino Herrera, Benjamín Costas, Luís E. C. Conceição, and Catarina I. M. Martins. 2013. Linking cortisol responsiveness and aggressive behaviour in gilthead seabream Sparus aurata: Indication of divergent coping styles. Applied Animal Behaviour Science 143: 75–81. https://doi.org/10.1016/j.applanim.2012.11.008.
25 Reviewed distribution maps for Gilthead seabream (Sparus aurata). 2016. Aquamaps.
26 Davis, P. S. 1988. Two occurrences of the gilthead, Sparus aurata Linnaeus 1758, on the coast of Northumberland, England. Journal of Fish Biology 33: 951–951. https://doi.org/10.1111/j.1095-8649.1988.tb05545.x.
27 Sánchez-Lamadrid, A. 2004. Effectiveness of releasing gilthead sea bream (Sparus aurata, L.) for stock enhancement in the bay of Cádiz. Aquaculture 231: 135–148. https://doi.org/10.1016/j.aquaculture.2003.08.015.
28 Craig, G., D. Paynter, I. Coscia, and S. Mariani. 2008. Settlement of gilthead sea bream Sparus aurata L. in a southern Irish Sea coastal habitat. Journal of Fish Biology 72: 287–291. https://doi.org/10.1111/j.1095-8649.2007.01644.x.
29 Ahmed, Mohamed S. 2011. Population dynamics and fisheries management of Gilthead sea bream, Sparus aurata (f. Sparidae) from Bardawil lagoon, North Sinai, Egypt. Egypt J. Aquat. Biol. & Fish. 15: 57–69.
30 Balik, Ismet, and Yilmaz Emre. 2013. Monthly variation in stock density and growth performance of juvenile gilthead seabream (Sparus aurata L., 1758) in Beymelek Lagoon, Antalya, Turkey. Pakistan J. Zool. 45: 687–693.
31 Mercier, Lény, Jacques Panfili, Christelle Paillon, Awa N’diaye, David Mouillot, and Audrey M. Darnaude. 2011. Otolith reading and multi-model inference for improved estimation of age and growth in the gilthead seabream Sparus aurata (L.). Estuarine, Coastal and Shelf Science 92: 534–545. https://doi.org/10.1016/j.ecss.2011.02.001.
32 Mercier, Lény, David Mouillot, Olivier Bruguier, Laurent Vigliola, and Audrey M. Darnaude. 2012. Multi-element otolith fingerprints unravel sea−lagoon lifetime migrations of gilthead sea bream Sparus aurata. Marine Ecology Progress Series 444: 175–194. https://doi.org/10.3354/meps09444.
33 Verdiell-Cubedo, David, Francisco J. Oliva-Paterna, Ana Ruiz-Navarro, and Mar Torralva. 2013. Assessing the nursery role for marine fish species in a hypersaline coastal lagoon (Mar Menor, Mediterranean Sea). Marine Biology Research 9: 739–748. https://doi.org/10.1080/17451000.2013.765580.
34 Chaoui, Lamya, Mohamed Hichem Kara, and Jean Pierre Quignard. 2006. Growth and reproduction of the gilthead seabream Sparus aurata in Mellah lagoon (north-eastern Algeria). Scientia Marina 70: 545–552. https://doi.org/10.3989/scimar.2006.70n3545.
35 Katselis, George, Constantin Koutsikopoulos, Evagelos Dimitriou, and Yiannis Rogdakis. 2003. Spatial patterns and temporal trends in the fisheries landings of the Messolonghi-Etoliko lagoons (Western Greek Coast). Scientia Marina 67. https://doi.org/10.3989/scimar.2003.67n4501.
36 Kraljević, Miro, and Jakov Dulčić. 1997. Age and growth of gilt-head sea bream (Sparus aurata L.) in the Mirna estuary, northern Adriatic. Fisheries Research 31: 249–255. https://doi.org/10.1016/S0165-7836(97)00016-7.
37 Guidetti, P., and S. Bussotti. 2002. Effects of seagrass canopy removal on fish in shallow Mediterranean seagrass (Cymodocea nodosa and Zostera noltii) meadows: a local-scale approach. Marine Biology 140: 445–453. https://doi.org/10.1007/s00227-001-0725-1.
38 Tancioni, L., S. Mariani, A. Maccaroni, A. Mariani, F. Massa, M. Scardi, and S. Cataudella. 2003. Locality-specific variation in the feeding of Sparus aurata L.: evidence from two Mediterranean lagoon systems. Estuarine, Coastal and Shelf Science 57: 469–474. https://doi.org/10.1016/S0272-7714(02)00376-1.
39 Franco, Anita, Piero Franzoi, Stefano Malavasi, Federico Riccato, Patrizia Torricelli, and Danilo Mainardi. 2006. Use of shallow water habitats by fish assemblages in a Mediterranean coastal lagoon. Estuarine, Coastal and Shelf Science 66: 67–83. https://doi.org/10.1016/j.ecss.2005.07.020.
40 Batzina, Alkisti, and Nafsika Karakatsouli. 2012. The presence of substrate as a means of environmental enrichment in intensively reared gilthead seabream Sparus aurata: Growth and behavioral effects. Aquaculture 370–371: 54–60. https://doi.org/10.1016/j.aquaculture.2012.10.005.
41 Froese, R., and D. Pauly. 2014. FishBase. World Wide Web electronic publication. www.fishbase.org.
42 FAO. 2014. The State of World Fisheries and Aquaculture 2014. Rome: Food and Agriculture Organization of the United Nations.
43 Watson, R., Jackie Alder, and Daniel Pauly. 2006. Fisheries for forage fish, 1950 to the present. In On the Multiple Uses of Forage Fish: from Ecosystems to Markets, ed. Jackie Alder and Daniel Pauly, 14:1–20. Fisheries Centre Research Reports 3. Vancouver, Canada: Fisheries Centre, University of British Columbia.
44 Mood, A. 2012. Average annual fish capture for species mostly used for fishmeal (2005-2009). fishcount.org.uk.
45 Mood, A., and P. Brooke. 2012. Estimating the Number of Farmed Fish Killed in Global Aquaculture Each Year.
46 Kopf, Von Kristin. 2012. Milliarden vs. Billionen: Große Zahlen. Sprachlog.
47 Weisstein, Eric W. 2018. Milliard. Text. MathWorld - a Wolfram Web resource. http://mathworld.wolfram.com/Milliard.html. Accessed February 2.
48 Ferrari, I., and A. R. Chieregato. 1981. Feeding habits of juvenile stages of Sparus auratus L., Dicentrarchus labrax L. and Mugilidae in a brackish embayment of the Po River Delta. Aquaculture 25: 243–257. https://doi.org/10.1016/0044-8486(81)90186-1.
49 Pita, C., S. Gamito, and K. Erzini. 2002. Feeding habits of the gilthead seabream (Sparus aurata) from the Ria Formosa (southern Portugal) as compared to the black seabream (Spondyliosoma cantharus) and the annular seabream (Diplodus annularis). Journal of Applied Ichthyology 18: 81–86. https://doi.org/10.1046/j.1439-0426.2002.00336.x.
50 Fernández-Diaz, C., E. Pascual, and M. Yúfera. 1994. Feeding behaviour and prey size selection of gilthead seabream, Sparus aurata, larvae fed on inert and live food. Marine Biology 118: 323–328. https://doi.org/10.1007/BF00349800.
51 Koven, W., Y. Barr, S. Lutzky, I. Ben-Atia, R. Weiss, M. Harel, P. Behrens, and A. Tandler. 2001. The effect of dietary arachidonic acid (20:4n−6) on growth, survival and resistance to handling stress in gilthead seabream (Sparus aurata) larvae. Aquaculture 193: 107–122. https://doi.org/10.1016/S0044-8486(00)00479-8.
52 Varela, J. L., I. Ruiz-Jarabo, L. Vargas-Chacoff, S. Arijo, J. M. León-Rubio, I. García-Millán, M. P. Martín del Río, M. A. Moriñigo, and J. M. Mancera. 2010. Dietary administration of probiotic Pdp11 promotes growth and improves stress tolerance to high stocking density in gilthead seabream Sparus auratus. Aquaculture 309: 265–271. https://doi.org/10.1016/j.aquaculture.2010.09.029.
53 Montero, D., L. Tort, L. Robaina, J. M. Vergara, and M. S. Izquierdo. 2001. Low vitamin E in diet reduces stress resistance of gilthead seabream (Sparus aurata) juveniles. Fish & Shellfish Immunology 11: 473–490. https://doi.org/10.1006/fsim.2000.0324.
54 Sarà, G., A. Oliveri, G. Martino, S. Serra, G. Meloni, and A. Pais. 2010. Response of captive seabass and seabream as behavioural indicator in aquaculture. Italian Journal of Animal Science 6: 823–825. https://doi.org/10.4081/ijas.2007.1s.823.
55 Montoya, A., J. F. López-Olmeda, M. Yúfera, M. J. Sánchez-Muros, and F. J. Sánchez-Vázquez. 2010. Feeding time synchronises daily rhythms of behaviour and digestive physiology in gilthead seabream (Sparus aurata). Aquaculture 306: 315–321. https://doi.org/10.1016/j.aquaculture.2010.06.023.
56 Sánchez, J. A., J. F. López-Olmeda, B. Blanco-Vives, and F. J. Sánchez-Vázquez. 2009. Effects of feeding schedule on locomotor activity rhythms and stress response in sea bream. Physiology & Behavior 98: 125–129. https://doi.org/10.1016/j.physbeh.2009.04.020.
57 Andrew, J. E, J Holm, S Kadri, and F. A Huntingford. 2004. The effect of competition on the feeding efficiency and feed handling behaviour in gilthead sea bream (Sparus aurata L.) held in tanks. Aquaculture 232: 317–331. https://doi.org/10.1016/S0044-8486(03)00528-3.
58 Cammarata, M., M. Vazzana, D. Accardi, and N. Parrinello. 2012. Seabream (Sparus aurata) long-term dominant-subordinate interplay affects phagocytosis by peritoneal cavity cells. Brain, Behavior, and Immunity 26: 580–587. https://doi.org/10.1016/j.bbi.2012.01.008.
59 Tort, L., J. Rotllant, C. Liarte, L. Acerete, A. Hernández, S. Ceulemans, P. Coutteau, and F. Padros. 2004. Effects of temperature decrease on feeding rates, immune indicators and histopathological changes of gilthead sea bream Sparus aurata fed with an experimental diet. Aquaculture 229: 55–65. https://doi.org/10.1016/S0044-8486(03)00403-4.
60 Ibarz, A., M. Beltrán, J. Fernández-Borràs, M. A. Gallardo, J. Sánchez, and J. Blasco. 2007. Alterations in lipid metabolism and use of energy depots of gilthead sea bream (Sparus aurata) at low temperatures. Aquaculture 262: 470–480. https://doi.org/10.1016/j.aquaculture.2006.11.008.
61 Katselis, George, Katerina Koukou, Evagelos Dimitriou, and Constantin Koutsikopoulos. 2007. Short-term seaward fish migration in the Messolonghi–Etoliko lagoons (Western Greek coast) in relation to climatic variables and the lunar cycle. Estuarine, Coastal and Shelf Science 73: 571–582. https://doi.org/10.1016/j.ecss.2007.02.010.
62 Saka, Sahin, Kürat Fırat, and Cüneyt Süzer. 2001. Effects Of Light Intensity On Early Life Development Of Gilthead Sea Bream Larvae (Sparus Aurata). Israeli Journal of Aquaculture - Bamidgeh 53: 139–146.
63 Tandler, Amos, and Sarah Helps. 1985. The effects of photoperiod and water exchange rate on growth and survival of gilthead sea bream (Sparus aurata, Linnaeus; Sparidae) from hatching to metamorphosis in mass rearing systems. Aquaculture 48: 71–82. https://doi.org/10.1016/0044-8486(85)90053-5.
64 Papoutsoglou, S. E., N. Karakatsouli, A. Batzina, E. S. Papoutsoglou, and A. Tsopelakos. 2008. Effect of music stimulus on gilthead seabream Sparus aurata physiology under different light intensity in a re-circulating water system. Journal of Fish Biology 73: 980–1004. https://doi.org/10.1111/j.1095-8649.2008.02001.x.
65 Kissil, George Wm, Ingrid Lupatsch, Abigail Elizur, and Yonathan Zohar. 2001. Long photoperiod delayed spawning and increased somatic growth in gilthead seabream (Sparus aurata). Aquaculture 200: 363–379. https://doi.org/10.1016/S0044-8486(01)00527-0.
66 Karakatsouli, Nafsika, Sofronios E. Papoutsoglou, Gianluca Pizzonia, Georgios Tsatsos, Aristeidis Tsopelakos, Stella Chadio, Dimitris Kalogiannis, Christina Dalla, Alexia Polissidis, and Zeta Papadopoulou-Daifoti. 2007. Effects of light spectrum on growth and physiological status of gilthead seabream Sparus aurata and rainbow trout Oncorhynchus mykiss reared under recirculating system conditions. Aquacultural Engineering 36: 302–309. https://doi.org/10.1016/j.aquaeng.2007.01.005.
67 Polo, A., M. Yúfera, and E. Pascual. 1991. Effects of temperature on egg and larval development of Sparus aurata L. Aquaculture 92: 367–375. https://doi.org/10.1016/0044-8486(91)90042-6.
68 Gallardo, M. Ángeles, Mónica Sala-Rabanal, Antoni Ibarz, Francesc Padrós, Josefina Blasco, Jaume Fernández-Borràs, and Josep Sánchez. 2003. Functional alterations associated with “winter syndrome” in gilthead sea bream (Sparus aurata). Aquaculture 223: 15–27. https://doi.org/10.1016/S0044-8486(03)00164-9.
69 Tort, L., F. Padrós, J. Rotllant, and S. Crespo. 1998. Winter syndrome in the gilthead sea breamSparus aurata. Immunological and histopathological features. Fish & Shellfish Immunology 8: 37–47. https://doi.org/10.1006/fsim.1997.0120.
70 Tort, L, J Rotllant, and L Rovira. 1998. Immunological suppression in gilthead sea bream Sparus aurata of the North-West Mediterranean at low temperatures. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 120: 175–179. https://doi.org/10.1016/S1095-6433(98)10027-2.
71 Rotllant, J., P. H. M. Balm, S. E. Wendelaar-Bonga, J. Pérez-Sánchez, and L. Tort. 2000. A drop in ambient temperature results in a transient reduction of interrenal ACTH responsiveness in the gilthead sea bream (Sparus aurata, L.). Fish Physiology and Biochemistry 23: 265–273. https://doi.org/10.1023/A:1007873811975.
72 Hernández, Juan M, Eucario Gasca-Leyva, Carmelo J León, and J. M Vergara. 2003. A growth model for gilthead seabream (Sparus aurata). Ecological Modelling 165: 265–283. https://doi.org/10.1016/S0304-3800(03)00095-4.
73 Bernabé, Gilbert. 1991. Acuicultura.
74 Bodinier, Charlotte, Elliott Sucré, Laura Lecurieux-Belfond, Eva Blondeau-Bidet, and Guy Charmantier. 2010. Ontogeny of osmoregulation and salinity tolerance in the gilthead sea bream Sparus aurata. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 157: 220–228. https://doi.org/10.1016/j.cbpa.2010.06.185.
75 Tandler, Amos, Fabio A. Anav, and Itzhak Choshniak. 1995. The effect of salinity on growth rate, survival and swimbladder inflation in gilthead seabream, Sparus aurata, larvae. Aquaculture 135: 343–353. https://doi.org/10.1016/0044-8486(95)01029-7.
76 Klaoudatos, S D, and A J Conides. 1996. Growth, food conversion, maintenance and long-term survival of gilthead sea bream, Sparus auratus L., juveniles after abrupt transfer to low salinity. Aquaculture Research 27: 765–774. https://doi.org/10.1046/j.1365-2109.1996.t01-1-00792.x.
77 Laiz-Carrión, Raúl, Susana Sangiao-Alvarellos, José M. Guzmán, María P. Martín del Río, José L. Soengas, and Juan M. Mancera. 2005. Growth performance of gilthead sea bream Sparus aurata in different osmotic conditions: Implications for osmoregulation and energy metabolism. Aquaculture 250: 849–861. https://doi.org/10.1016/j.aquaculture.2005.05.021.
78 Riera, Rodrigo, Víctor M. Tuset, Myriam Rodríguez, Óscar Monterroso, and Antoni Lombarte. 2017. Analyzing functional diversity to determine the effects of fish cages in insular coastal wild fish assemblages. Aquaculture 479: 384–395. https://doi.org/10.1016/j.aquaculture.2017.06.014.
79 Steinhausen, Maria Faldborg, John Fleng Steffensen, and Niels Gerner Andersen. 2010. The effects of swimming pattern on the energy use of gilthead seabream (Sparus aurata L.). Marine and Freshwater Behaviour and Physiology 43: 227–241. https://doi.org/10.1080/10236244.2010.501135.
80 Mariani, Stefano. 2006. Life-history- and ecosystem-driven variation in composition and residence pattern of seabream species (Perciformes: Sparidae) in two Mediterranean coastal lagoons. Marine Pollution Bulletin 53. Recent Developments in Estuarine Ecology and Management: 121–127. https://doi.org/10.1016/j.marpolbul.2005.09.019.
81 Zohar, Yonathan, M. Harel, S. Hassin, and Amos Tandler. 1995. Gilt-head sea bream (Sparus aurata). In Broodstock Management and Egg and Larval Quality, ed. Niall R. Bromage and Ronald J. Roberts, 94–117. Wiley.
82 Colloca, F., and S. Cerasi. 2005. Cultured Aquatic Species Information Programme. Sparus aurata. Rome: FAO Fisheries and Aquaculture Department.
83 Brown, Richard Cameron. 2003. Genetic Management and Selective Breeding in Farmed Populations of Gilthead Seabream (Sparus aurata). Thesis or Dissertation, University of Stirling.
84 Ferraresso, Serena, Nicola Vitulo, Alba N. Mininni, Chiara Romualdi, Barbara Cardazzo, Enrico Negrisolo, Richard Reinhardt, Adelino VM Canario, Tomaso Patarnello, and Luca Bargelloni. 2008. Development and validation of a gene expression oligo microarray for the gilthead sea bream (Sparus aurata). BMC Genomics 9: 580. https://doi.org/10.1186/1471-2164-9-580.
85 Andrades, J. A., J. Becerra, and P. Fernández-Llebrez. 1996. Skeletal deformities in larval, juvenile and adult stages of cultured gilthead sea bream (Sparus aurata L.). Aquaculture 141: 1–11. https://doi.org/10.1016/0044-8486(95)01226-5.
86 Faustino, M., and D. M. Power. 2001. Osteologic development of the viscerocranial skeleton in sea bream: alternative ossification strategies in teleost fish. Journal of Fish Biology 58: 537–572. https://doi.org/10.1111/j.1095-8649.2001.tb02272.x.
87 Elbal, M. T, M. P Garcı́a Hernández, M. T Lozano, and B Agulleiro. 2004. Development of the digestive tract of gilthead sea bream (Sparus aurata L.). Light and electron microscopic studies. Aquaculture 234: 215–238. https://doi.org/10.1016/j.aquaculture.2003.11.028.
88 Russo, T., C. Costa, and S. Cataudella. 2007. Correspondence between shape and feeding habit changes throughout ontogeny of gilthead sea bream Sparus aurata L., 1758. Journal of Fish Biology 71: 629–656. https://doi.org/10.1111/j.1095-8649.2007.01528.x.
89 Parra, G, and M Yúfera. 2000. Feeding, physiology and growth responses in first-feeding gilthead seabream (Sparus aurata L.) larvae in relation to prey density. Journal of Experimental Marine Biology and Ecology 243: 1–15. https://doi.org/10.1016/S0022-0981(99)00106-9.
90 Pawson, M.G., and G.D. Pickett. 1996. The Annual Pattern of Condition and Maturity in Bass, Dicentrarchus Labrax, in Waters Around England and Wales. Journal of the Marine Biological Association of the United Kingdom 76: 107. https://doi.org/10.1017/S0025315400029040.
91 Brusléa-Sicard, S., and B. Fourcault. 1997. Recognition of sex-inverting protandric Sparus aurata: ultrastructural aspects. Journal of Fish Biology 50: 1094–1103. https://doi.org/10.1111/j.1095-8649.1997.tb01633.x.
92 Mehanna, Sahar Fahmy. 2007. A preliminary assessment and management of gilthead bream Sparus aurata in the Port Said fishery, the Southeastern Mediterranean, Egypt. Turkish Journal of Fisheries and Aquatic Sciences 7: 123–130.
93 Kadmon, G., Z. Yaron, and H. Gordin. 1985. Sequence of gonadal events and oestradiol levels in Sparus aurata (L.) under two photoperiod regimes. Journal of Fish Biology 26: 609–620. https://doi.org/10.1111/j.1095-8649.1985.tb04301.x.
94 Wassef, Elham A. 1990. Growth rate of gilthead bream Sparus aurata L. J. K. A. U. Mar. Sci. 1: 55–65.
95 Gonçalves, J. M. S., L. Bentes, P. G. Lino, J. Ribeiro, A. V. M. Canário, and K. Erzini. 1997. Weight-length relationships for selected fish species of the small-scale demersal fisheries of the south and south-west coast of Portugal. Fisheries Research 30: 253–256. https://doi.org/10.1016/S0165-7836(96)00569-3.
96 Zohar, Y., M. Abraham, and H. Gordin. 1978. The gonadal cycle of the captivity-reared hermaphroditic teleost Sparus aurata (L.) during the first two years of life. Annales de Biologie Animale Biochimie Biophysique 18: 877–882. https://doi.org/10.1051/rnd:19780519.
97 Wong, Ten-Tsao, Shigeho Ijiri, and Yonathan Zohar. 2006. Molecular Biology of Ovarian Aromatase in Sex Reversal: Complementary DNA and 5′-Flanking Region Isolation and Differential Expression of Ovarian Aromatase in the Gilthead Seabream (Sparus aurata). Biology of Reproduction 74: 857–864. https://doi.org/10.1095/biolreprod.105.045351.
98 Navarro, Ana, María J. Zamorano, Silvia Hildebrandt, Rafael Ginés, Cristóbal Aguilera, and Juan M. Afonso. 2009. Estimates of heritabilities and genetic correlations for growth and carcass traits in gilthead seabream (Sparus auratus L.), under industrial conditions. Aquaculture 289: 225–230. https://doi.org/10.1016/j.aquaculture.2008.12.024.
99 Addis, Maria Filippa, Roberto Cappuccinelli, Vittorio Tedde, Daniela Pagnozzi, Maria Cristina Porcu, Elia Bonaglini, Tonina Roggio, and Sergio Uzzau. 2010. Proteomic analysis of muscle tissue from gilthead sea bream (Sparus aurata, L.) farmed in offshore floating cages. Aquaculture 309: 245–252. https://doi.org/10.1016/j.aquaculture.2010.08.022.
100 Boglione, Clara, Flavio Gagliardi, Michele Scardi, and Stefano Cataudella. 2001. Skeletal descriptors and quality assessment in larvae and post-larvae of wild-caught and hatchery-reared gilthead sea bream (Sparus aurata L. 1758). Aquaculture 192: 1–22. https://doi.org/10.1016/S0044-8486(00)00446-4.
101 Carrillo, J, G Koumoundouros, P Divanach, and J Martinez. 2001. Morphological malformations of the lateral line in reared gilthead sea bream (Sparus aurata L. 1758). Aquaculture 192: 281–290. https://doi.org/10.1016/S0044-8486(00)00454-3.
102 Paperna, I. 1978. Swimbladder and skeletal deformations in hatchery bred Spams aurata. Journal of Fish Biology 12: 109–114. https://doi.org/10.1111/j.1095-8649.1978.tb04157.x.
103 Koumoundouros, G., G. Oran, P. Divanach, S. Stefanakis, and M. Kentouri. 1997. The opercular complex deformity in intensive gilthead sea bream (Sparus aurata L.) larviculture. Moment of apparition and description. Aquaculture 156: 165–177. https://doi.org/10.1016/S0044-8486(97)89294-0.
104 Afonso, J. M., D. Montero, L. Robaina, N. Astorga, M. S. Izquierdo, and R. Ginés. 2000. Association of a lordosis-scoliosis-kyphosis deformity in gilthead seabream (Sparus aurata) with family structure. Fish Physiology and Biochemistry 22: 159–163. https://doi.org/10.1023/A:1007811702624.
105 Castro, Jaime, Ania Pino-Querido, Miguel Hermida, David Chavarrías, Roberto Romero, Luis A. García-Cortés, Miguel A. Toro, and Paulino Martínez. 2008. Heritability of skeleton abnormalities (lordosis, lack of operculum) in gilthead seabream (Sparus aurata) supported by microsatellite family data. Aquaculture 279: 18–22. https://doi.org/10.1016/j.aquaculture.2008.04.023.
106 Balebona, M. C., M. A. Morinigo, J. A. Andrades, J. A. Santamaria, J. Becerra, and J. S. Borrego. 1993. Microbiological study of gilthead sea bream S. aurata L. affected by lordosis a skeletal deformity. Bull. Eur. Assoc. Fish Pathol. 13: 33.
107 Chatain, Beatrice. 1994. Abnormal swimbladder development and lordosis in sea bass (Dicentrarchus labrax) and sea bream (Sparus auratus). Aquaculture 119: 371–379. https://doi.org/0044-8486/94.
108 Reyes-Tomassini, Jose J. 2009. Behavioral and Neuroendocrine Correlates of Sex Change in the Gilthead Seabream (Sparus aurata). University of Maryland.
109 Gordin, H., and Y. Zohar. 1978. Induced spawning of Sparus aurata (L.) by means of hormonal treatments. Annales de Biologie Animale Biochimie Biophysique 18: 985–990. https://doi.org/10.1051/rnd:19780535.
110 Liarte, Sergio, Elena Chaves-Pozo, Alicia García-Alcazar, Victoriano Mulero, José Meseguer, and Alfonsa García-Ayala. 2007. Testicular involution prior to sex change in gilthead seabream is characterized by a decrease in DMRT1 gene expression and by massive leukocyte infiltration. Reproductive Biology and Endocrinology 5: 20. https://doi.org/10.1186/1477-7827-5-20.
111 Barbaro, A., A. Francescon, G. Bozzato, A. Merlin, P. Belvedere, and L. Colombo. 1997. Induction of spawning in gilthead seabream, Sparus aurata L., by a long-acting GnRH agonist and its effects on egg quality and daily timing of spawning. Aquaculture 154: 349–359. https://doi.org/10.1016/S0044-8486(97)00067-7.
112 Gorshkov, S., H. Gordin, G. Gorshkova, and W. Knibb. 1997. Reproductive constraints for family selection of the gilthead seabream (Sparus aurata L.). Israeli Journal of Aquaculture 49: 124–134. CABDirect2.
113 Meiri, Iris, Yoav Gothilf, Yonathan Zohar, and Abigail Elizur. 2002. Physiological changes in the spawning gilthead seabream, Sparus aurata, succeeding the removal of males. Journal of Experimental Zoology 292: 555–564. https://doi.org/10.1002/jez.10072.
114 Zohar, Y., and H. Gordin. 1979. Spawning kinetics in the gilthead sea-bream, Sparus aurata L. after low doses of human chronic gonadotropin. Journal of Fish Biology 15: 665–670. https://doi.org/10.1111/j.1095-8649.1979.tb03675.x.
115 Saavedra, Margarida, and Pedro Pousão-Ferreira. 2006. A preliminary study on the effect of lunar cycles on the spawning behaviour of the gilt-head sea bream, Sparus aurata. Journal of the Marine Biological Association of the United Kingdom 86: 899–901. https://doi.org/10.1017/S0025315406013841.
116 Zohar, Y., A. Goren, M. Tosky, G. Pagelson, D. Leibovitz, and Y. Koch. 1989. The bioactivity of gonadotropin releasing hormones and its regulation in the gilthead seabream,Sparus aurata: in vivo andin vitro studies. Fish Physiology and Biochemistry 7: 59–67. https://doi.org/10.1007/BF00004690.
117 Lasserre, G., and P. J. Labourg. 1974. Comparison of growth of Sparus auratus L. in regions of Arcachon and Sete (2nd note). Vie et Milieu Serie A - Biologie Marine 24: 357–363.
118 Kolkovski, S., A. Arieli, and A. Tandler. 1997. Visual and chemical cues stimulate microdiet ingestion in sea bream larvae. Aquaculture International 5: 527–536. https://doi.org/10.1023/A:1018305416501.
119 Hubbard, P. C., E. N. Barata, and A. V. M. Canário. 2003. Olfactory Sensitivity to Catecholamines and their Metabolites in the Goldfish. Chemical Senses 28: 207–218. https://doi.org/10.1093/chemse/28.3.207.
120 Brown, Culum. 2015. Fish intelligence, sentience and ethics. Animal Cognition 18: 1–17. https://doi.org/10.1007/s10071-014-0761-0.
121 Amundsen, Lasse, and Martin Landro. 2011. Marine seismic sources part VIII: Fish hear a great deal. Recent Advances in Technology 8: 1–5.
122 Buscaino, Giuseppa, Francesco Filiciotto, Gaspare Buffa, Antonio Bellante, Vincenzo Di Stefano, Anna Assenza, Francesco Fazio, Giovanni Caola, and Salvatore Mazzola. 2010. Impact of an acoustic stimulus on the motility and blood parameters of European sea bass (Dicentrarchus labrax L.) and gilthead sea bream (Sparus aurata L.). Marine Environmental Research 69: 136–142. https://doi.org/10.1016/j.marenvres.2009.09.004.
123 Filiciotto, Francesco, Vincenzo Maximiliano Giacalone, Francesco Fazio, Gaspare Buffa, Giuseppe Piccione, Vincenzo Maccarrone, Vincenzo Di Stefano, Salvatore Mazzola, and Giuseppa Buscaino. 2013. Effect of acoustic environment on gilthead sea bream (Sparus aurata): Sea and onshore aquaculture background noise. Aquaculture 414–415: 36–45. https://doi.org/10.1016/j.aquaculture.2013.07.042.
124 Dijkgraaf, S. 1967. Biological significance of the lateral line organs. In Lateral Line Detectors: Proceedings. Edited by Phyllis H. Cahn, ed. Phyllis H. Cahn, 83–95. Indiana University Press.
125 Bleckmann, Horst. 1986. Role of the Lateral Line in Fish Behaviour. In The Behaviour of Teleost Fishes, ed. Tony J. Pitcher, 177–202. Springer US.
126 Suckling, J. A. 1967. Trunk lateral line nerves: some anatomical aspects. In Lateral Line Detectors: Proceedings., ed. Phyllis H. Cahn, 45–52. Indiana University Press.
127 Bauchot, M.-L., and J.-C. Hureau. 1986. Sparidae. In Fishes of the North-eastern Atlantic and the Mediterranean (FNAM). P.J.P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen and E. Tortonese (eds)., 2:883–907. Unesco, Paris.
128 Bagni, M., C. Civitareale, A. Priori, A. Ballerini, M. Finoia, G. Brambilla, and G. Marino. 2007. Pre-slaughter crowding stress and killing procedures affecting quality and welfare in sea bass (Dicentrarchus labrax) and sea bream (Sparus aurata). Aquaculture 263: 52–60. https://doi.org/10.1016/j.aquaculture.2006.07.049.
129 Tort, L., J. O. Sunyer, E. Gómez, and A. Molinero. 1996. Crowding stress induces changes in serum haemolytic and agglutinating activity in the gilthead sea bream Sparus aurata. Veterinary Immunology and Immunopathology 51: 179–188. https://doi.org/10.1016/0165-2427(95)05502-9.
130 Arends, R. J., J. M. Mancera, J. L. Munoz, SE Wendelaar Bonga, and G. Flik. 1999. The stress response of the gilthead sea bream (Sparus aurata L.) to air exposure and confinement. Journal of Endocrinology 163: 149–157. https://doi.org/10.1677/joe.0.1630149.
131 Montero, D., M. S. Izquierdo, L. Tort, L. Robaina, and J. M. Vergara. 1999. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiology and Biochemistry 20: 53–60. https://doi.org/10.1023/A:1007719928905.
132 Rotllant, J., P. H. M. Balm, J. Pérez-Sánchez, S. E. Wendelaar-Bonga, and L. Tort. 2001. Pituitary and Interrenal Function in Gilthead Sea Bream (Sparus aurata L., Teleostei) after Handling and Confinement Stress. General and Comparative Endocrinology 121: 333–342. https://doi.org/10.1006/gcen.2001.7604.
133 Ortuño, J., M. A. Esteban, and J. Meseguer. 2001. Effects of short-term crowding stress on the gilthead seabream (Sparus aurata L.) innate immune response. Fish & Shellfish Immunology 11: 187–197. https://doi.org/10.1006/fsim.2000.0304.
134 Matos, Elisabete, Amparo Gonçalves, Maria Leonor Nunes, Maria Teresa Dinis, and Jorge Dias. 2010. Effect of harvesting stress and slaughter conditions on selected flesh quality criteria of gilthead seabream (Sparus aurata). Aquaculture 305: 66–72. https://doi.org/10.1016/j.aquaculture.2010.04.020.
135 Goldan, O. 1992. The control of growth depensation in juvenile gilthead sea bream Sparus aurata. Dissertation, The Hebrew University of Jerusalem.
136 Réale, Denis, Simon M. Reader, Daniel Sol, Peter T. McDougall, and Niels J. Dingemanse. 2007. Integrating animal temperament within ecology and evolution. Biological Reviews 82: 291–318. https://doi.org/10.1111/j.1469-185X.2007.00010.x.
137 Robb, D H F, and S C Kestin. 2002. Methods Used to Kill Fish: Field Observations and Literature Reviewed. Animal Welfare 11: 269–282.
138 van De Vis, Hans, Steve Kestin, David Robb, Jörg Oehlenschläger, Bert Lambooij, Werner Münkner, Holmer Kuhlmann, et al. 2003. Is humane slaughter of fish possible for industry? Aquaculture Research 34: 211–220. https://doi.org/10.1046/j.1365-2109.2003.00804.x.





. Computer generated distribution maps for Sparus aurata (Gilthead seabream), with modelled year 2050 native range map based on IPCC RCP8.5 emissions scenario. Retrieved from https://www.aquamaps.org.")





Probably, we updated the profile. Check the version number in the head of the page. For more information on the version, see the FAQ about this. Why do we update profiles? Not just do we want to include new research that has come out, but we are continuously developing the database itself. For example, we changed the structure of entries in criteria or we added explanations for scores in the WelfareCheck | farm. And we are always refining our scoring rules.
The centre of the Overview is an array of criteria covering basic features and behaviours of the species. Each of this information comes from our literature search on the species. If we researched a full Dossier on the species, probably all criteria in the Overview will be covered and thus filled. This was our way to go when we first set up the database.
Because Dossiers are time consuming to research, we switched to focusing on WelfareChecks. These are much shorter profiles covering just 10 criteria we deemed important when it comes to behaviour and welfare in aquaculture (and lately fisheries, too). Also, WelfareChecks contain the assessment of the welfare potential of a species which has become the main feature of the fair-fish database over time. Because WelfareChecks do not cover as many criteria as a Dossier, we don't have the information to fill all blanks in the Overview, as this information is "not investigated by us yet".
Our long-term goal is to go back to researching Dossiers for all species covered in the fair-fish database once we set up WelfareChecks for each of them. If you would like to support us financially with this, please get in touch at ffdb@fair-fish.net
See the question "What does "not investigated by us yet" mean?". In short, if we have not had a look in the literature - or in other words, if we have not investigated a criterion - we cannot know the data. If we have already checked the literature on a criterion and could not find anything, it is "no data found yet". You spotted a "no data found yet" where you know data exists? Get in touch with us at ffdb@fair-fish.net!
Once you have clicked on "show details", the entry for a criterion will unfold and display the summarised information we collected from the scientific literature – complete with the reference(s).
As reference style we chose "Springer Humanities (numeric, brackets)" which presents itself in the database as a number in a grey box. Mouse over the box to see the reference; click on it to jump to the bibliography at the bottom of the page. But what does "[x]-[y]" refer to?
This is the way we mark secondary citations. In this case, we read reference "y", but not reference "x", and cite "x" as mentioned in "y". We try to avoid citing secondary references as best as possible and instead read the original source ourselves. Sometimes we have to resort to citing secondarily, though, when the original source is: a) very old or not (digitally) available for other reasons, b) in a language no one in the team understands. Seldomly, it also happens that we are running out of time on a profile and cannot afford to read the original. As mentioned, though, we try to avoid it, as citing mistakes may always happen (and we don't want to copy the mistake) and as misunderstandings may occur by interpreting the secondarily cited information incorrectly.
If you spot a secondary reference and would like to send us the original work, please contact us at ffdb@fair-fish.net
In general, we aim at giving a good representation of the literature published on the respective species and read as much as we can. We do have a time budget on each profile, though. This is around 80-100 hours for a WelfareCheck and around 300 hours for a Dossier. It might thus be that we simply did not come around to reading the paper.
It is also possible, though, that we did have to make a decision between several papers on the same topic. If there are too many papers on one issue than we manage to read in time, we have to select a sample. On certain topics that currently attract a lot of attention, it might be beneficial to opt for the more recent papers; on other topics, especially in basic research on behaviour in the wild, the older papers might be the go-to source.
And speaking of time: the paper you are missing from the profile might have come out after the profile was published. For the publication date, please check the head of the profile at "cite this profile". We currently update profiles every 6-7 years.
If your paper slipped through the cracks and you would like us to consider it, please get in touch at ffdb@fair-fish.net
This number, for example "C | 2.1 (2022-11-02)", contains 4 parts:
- "C" marks the appearance – the design level – of the profile part. In WelfareChecks | farm, appearance "C" is our most recent one with consistent age class and label (WILD, FARM, LAB) structure across all criteria.
- "2." marks the number of major releases within this appearance. Here, it is major release 2. Major releases include e.g. changes of the WelfareScore. Even if we just add one paper – if it changes the score for one or several criteria, we will mark this as a major update for the profile. With a change to a new appearance, the major release will be re-set to 1.
- ".1" marks the number of minor updates within this appearance. Here, it is minor update 1. With minor updates, we mean changes in formatting, grammar, orthography. It can also mean adding new papers, but if these papers only confirm the score and don't change it, it will be "minor" in our book. With a change to a new appearance, the minor update will be re-set to 0.
- "(2022-11-02)" is the date of the last change – be it the initial release of the part, a minor, or a major update. The nature of the changes you may find out in the changelog next to the version number.
If an Advice, for example, has an initial release date and then just a minor update date due to link corrections, it means that – apart from correcting links – the Advice has not been updated in a major way since its initial release. Please take this into account when consulting any part of the database.
First up, you will find answers to questions for the specific page you are on. Scrolling down in the FAQ window, there are also answers to more general questions. Explore our website and the other sub pages and find there the answers to questions relevant for those pages.
In the fair-fish database, when you have chosen a species (either by searching in the search bar or in the species tree), the landing page is an Overview, introducing the most important information to know about the species that we have come across during our literatures search, including common names, images, distribution, habitat and growth characteristics, swimming aspects, reproduction, social behaviour but also handling details. To dive deeper, visit the Dossier where we collect all available ethological findings (and more) on the most important aspects during the life course, both biologically and concerning the habitat. In contrast to the Overview, we present the findings in more detail citing the scientific references.
Depending on whether the species is farmed or wild caught, you will be interested in different branches of the database.
Farm branch
Founded in 2013, the farm branch of the fair-fish database focuses on farmed aquatic species.
Catch branch
Founded in 2022, the catch branch of the fair-fish database focuses on wild-caught aquatic species.
The heart of the farm branch of the fair-fish database is the welfare assessment – or WelfareCheck | farm – resulting in the WelfareScore | farm for each species. The WelfareCheck | farm is a condensed assessment of the species' likelihood and potential for good welfare in aquaculture, based on welfare-related findings for 10 crucial criteria (home range, depth range, migration, reproduction, aggregation, aggression, substrate, stress, malformations, slaughter).
For those species with a Dossier, we conclude to-be-preferred farming conditions in the Advice | farm. They are not meant to be as detailed as a rearing manual but instead, challenge current farming standards and often take the form of what not to do.
In parallel to farm, the main element of the catch branch of the fair-fish database is the welfare assessment – or WelfareCheck | catch – with the WelfareScore | catch for each species caught with a specific catching method. The WelfareCheck | catch, too, is a condensed assessment of the species' likelihood and potential for good welfare – or better yet avoidance of decrease of good welfare – this time in fisheries. We base this on findings on welfare hazards in 10 steps along the catching process (prospection, setting, catching, emersion, release from gear, bycatch avoidance, sorting, discarding, storing, slaughter).
In contrast to the farm profiles, in the catch branch we assess the welfare separately for each method that the focus species is caught with. In the case of a species exclusively caught with one method, there will be one WelfareCheck, whereas in other species, there will be as many WelfareChecks as there are methods to catch the species with.
Summarising our findings of all WelfareChecks | catch for one species in Advice | catch, we conclude which catching method is the least welfare threatening for this species and which changes to the gear or the catching process will potentially result in improvements of welfare.
Welfare of aquatic species is at the heart of the fair-fish database. In our definition of welfare, we follow Broom (1986): “The welfare of an individual is its state as regards its attempts to cope with its environment.” Thus, welfare may be perceived as a continuum on which an individual rates “good” or “poor” or everything in between.
We pursue what could be called a combination of not only a) valuing the freedom from injuries and stress (function-based approach) but b) supporting attempts to provide rewarding experiences and cognitive challenges (feelings-based approach) as well as c) arguing for enclosures that mimic the wild habitat as best as possible and allow for natural behaviour (nature-based approach).
Try mousing over the element you are interested in - oftentimes you will find explanations this way. If not, there will be FAQ on many of the sub-pages with answers to questions that apply to the respective sub-page. If your question is not among those, contact us at ffdb@fair-fish.net.
It's right here! We decided to re-name it to fair-fish database for several reasons. The database has grown beyond dealing purely with ethology, more towards welfare in general – and so much more. Also, the partners fair-fish and FishEthoGroup decided to re-organise their partnership. While maintaining our friendship, we also desire for greater independence. So, the name "fair-fish database" establishes it as a fair-fish endeavour.